作者:MANUEL D. VALDEARENAS MARTIN
視覺系統是光刺激的特異性受體,不僅會作用於動物:讓我們回想起由葉綠素介導的植物中的光合作用,其將二氧化碳和水轉化為碳水化合物和氧氣,並積累 的能量。
光能可以四種不同方式作用於生物體:影響代謝過程,影響運動,與色素沉著有關或引起視覺本身(物理,化學,神經或心理機制)。
1.光對代謝的影響。
晝夜的每日節奏以及該節奏的季節性變化都會導致整個動物規模的光週期,甚至影響性週期。
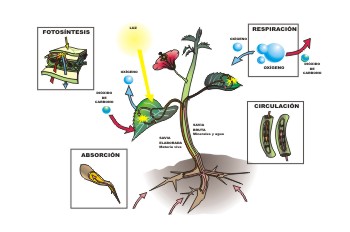
圖1,光與植物新陳代謝
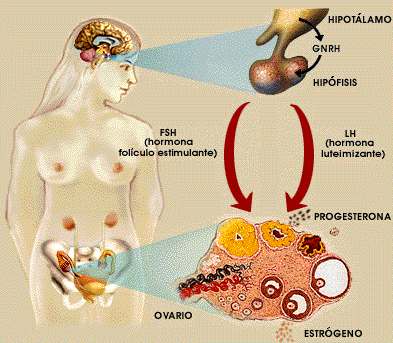
圖2,女性性週期
2.光線對運動的影響。
它們分為三種不同的類型:
- 對光
趨光性是植物的典型特徵,由無柄器官朝著光或遠離光的運動組成(正向,負向或橫向向光性)。

圖3,向日葵的植物向光性
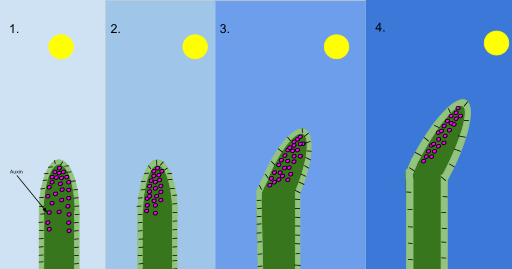
圖4,向光性機理(生長素
- 光電耦合
沒有方向性的運動改變。 矯正運動(運動加速或減慢)和斜運動學(運動朝著或遠離光的偏離)之間有區別。
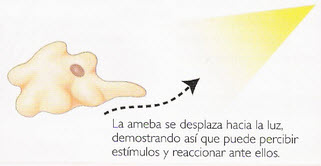
圖5,變形蟲在阿米巴中的運動
- 光軸
方向性改變運動性。 它需要能夠區分光源方向的接收器。

圖6,飛蛾的趨光性
趨光性分為:
觸軸
這需要存在一個受體器官,該器官比較連續刺激的強度,以不規則的方式(例如鞭毛的方向)相對於光引導位移。
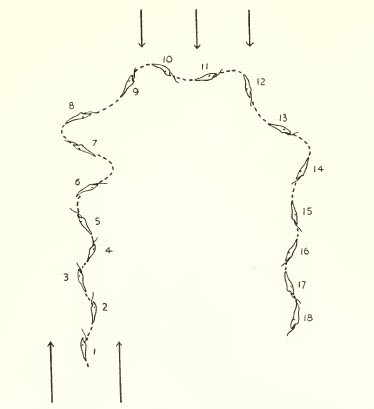
圖7,在裸藻中的氣候軸
對流
它需要兩個對稱的接收器,並且通過比較每個接收器的刺激強度(例如蠕蟲的位移)來進行更精確的移動。
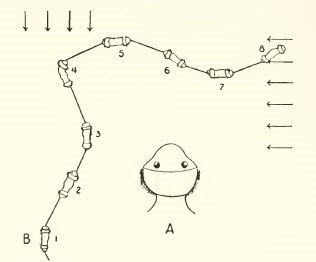
圖8,蠕蟲的對光軸
四軸
定向直接朝向或來自光源。 儘管單個受體就足夠了,但它必須是複雜的並且由多個元素組成。 另外,必須有一個中樞神經組織來分析刺激並抑制不必要的或強度較小的刺激(昆蟲的複眼)。
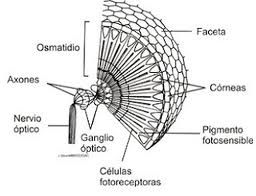
圖9,複眼
中軸
它不僅具有移入或移出光線的能力,而且還可以以一定角度移動(光線羅盤反應)。 鳥類的導航系統以及偏振第二光(蜜蜂的舞動)的可能方向都基於此。 它還包括圖像前面的方向以及有助於空氣或水導航的背側或腹側光反應。
圖10,鳥的menotaxis
趨動性
其中包括與當前刺激進行比較的視覺記憶。
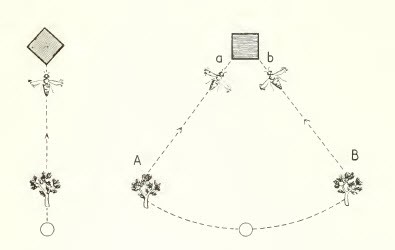
圖11,蜜蜂的驅蟲
3.光對色素沉著的影響。
存在的吸收光或過濾光的顏料,具有較少的保護(隱性)或示範性(扇形)功能,具有社會或生殖價值。

圖12,鞋底,色素沉著的保護功能

圖13,孔雀色素沉著的功能
它需要有組織的細胞或細胞組的存在:色譜。色素可以是黑色素(黑色素),它將由脂溶性類胡蘿蔔素(脂質體)或鳥嘌呤(胍基)組成,最後兩種具有很大的色差和虹彩。

圖14,棕褐色(黑色素)

圖15,變色龍(guanophores)
光對顏料的作用可以分為三種類型:
直接在皮膚的黑色素細胞上,甚至在視網膜或虹膜外胚層上:在魚類和兩棲類頭足類動物中直接瞳孔收縮。
通過眼睛暗示。
間接通過中腦(某些魚)或鬆果體(扁桃,硬骨魚)。
4.這樣的願景。
視覺的基本和原始功能是控制運動,以盡可能輕鬆地獲得最佳環境。這用於避免障礙,追捕獵物或從掠食者逃走,並且在人類中需要眼睛與前庭裝置之間的緊密關係。

圖16,猩猩的手眼協調
從本質上講,較高水平的視覺功能是有意識的,儘管在較低級的動物(例如,螞蟻的步態)中它們不是,而且在控制論反饋機制調節的機器中也不自然。視覺的“意識”過程與終端站從基核到皮層的遷移相吻合,或者已經歸因於此(這是進化史上的最新事實)。
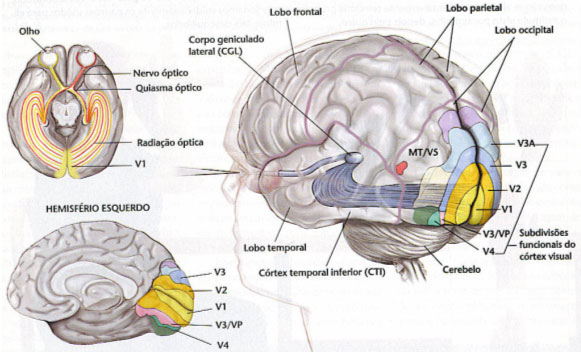
圖17,人的視覺通路:視覺皮層化
視覺裝置的演變
儘管在較低等的動物中,整個外胚層都有漫射的光敏性,但很快它的一部分專門研究僅限於感光器的光接收。它們的特徵在於存在顏料:化學惰性的黑色素和僅作為吸收劑的黑色素,以及可見的顏料本身。所有這些都是基於與蛋白質相關或不相關的類胡蘿蔔素,除了葉綠素以外,它們還存在於植物界(葉黃素)中。當達到具有光學成像能力的眼睛水平時,所有色素都與維生素A(A1或A2)特別相關。
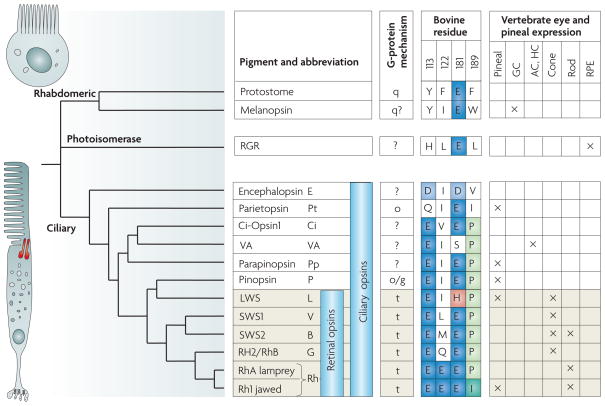
圖18,感光體和顏料的演變
在單細胞生物中,漫射光敏感性可以集中在與纖毛或鞭毛相關的點(運動功能)。在鞭毛蟲中,這種“視點”或“柱頭”具有色素,有時還具有可伸縮結構,該結構會聚光並像透鏡(晶體)一樣起作用。
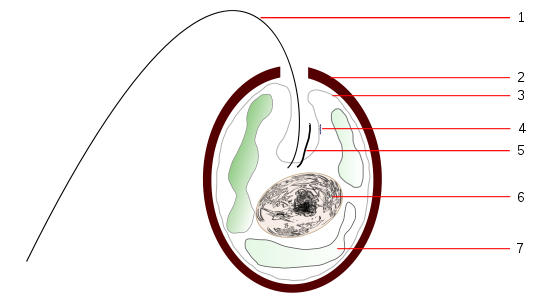
圖19,曲氯崙方案
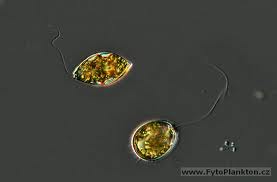
圖20,氣管單胞菌
在多細胞生物中,一些外胚層細胞很快就專門從事光接收,如雙極細胞(遠端接收端和近端發射端)或橢圓形的非極性細胞。從這裡開始無脊椎動物的眼睛進化。
无脊椎动物的眼睛
它源自表面外胚层,其次与神经系统连接。
它可以是简单的,可以是单细胞或多细胞的,也可以是复合的。
-简单的眼睛。
它由一个或多个不具有功能关联的光敏细胞组成。
当它是多细胞的时,它仍然可以是上皮下的(仅仅是光敏细胞的集合),或者在更晚期阶段是上皮的上空。这最后一种类型穿过扁平的杯状(暗室的形成),最后穿过水泡阶段,这在头足类动物的眼中仍是完美的,在该类动物中,上皮的增厚已经形成晶状体的“阴影”,并且收缩的学生

图21,紫草
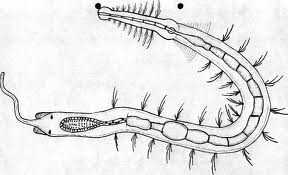
图22,Stilaria L解剖图
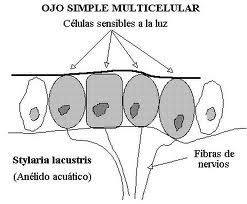
图23,平眼,紫丁香。

图24,骨菌。
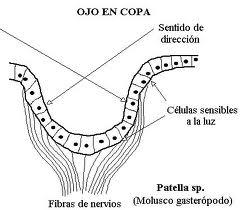
图25,eye骨SP杯中的眼睛

图26,螺旋果皮
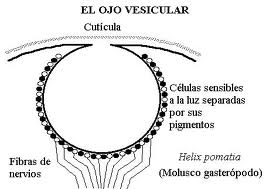
图27,螺旋P中的囊泡眼
尽管脊椎动物存在着倒置的视网膜(其中感光元件位于最深的一层,而递质位于表面,这可以使前者获得更好的营养),但是某些软体动物和蜘蛛具有这种特征新的视网膜组织,而不是“浇注”或直接视网膜。视网膜方向的这种变化是根据连续进化理论来解释的最困难的过程之一。此外,在必须生活在不良光照条件下的动物中,视网膜后面会出现反射性晶体层或绒毡层。
-复眼。
在复眼中,感觉元件在结构上和感觉上分组。它们中的每一个都被称为Ommatidium,由角膜小面,晶状体,虹膜色素细胞和视网膜组成,这些细胞由感光细胞组成,它们的延伸以它们中的几个称为横纹肌的共同延长为起点,在横纹中开始传播对神经中枢的冲动。
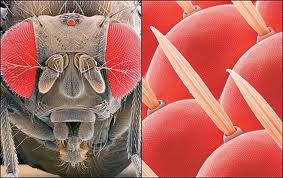
图28,复眼
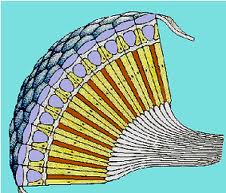
图29,复眼方案
节肢动物特有的复眼在蜘蛛,甲壳类和昆虫中采用不同的品种。尽管只能使视敏度降低,视敏度随横纹肌上视网膜浓度的高低而变化,但复眼具有分析光偏振面的能力,这是空间定向的基本功能。
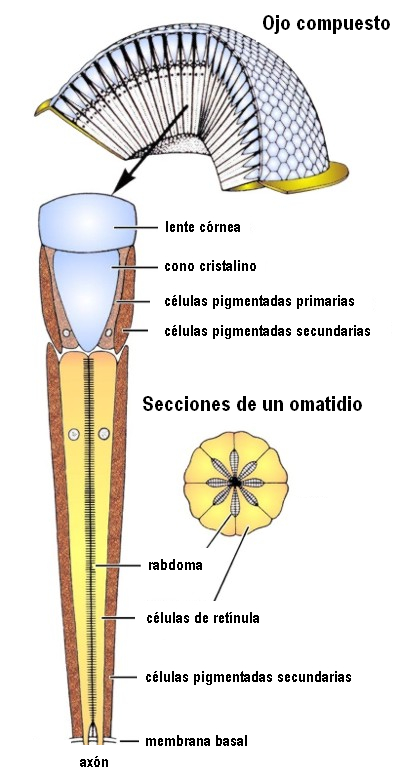
图30,mm孔的结构
脊椎动物的眼睛
它假设是一场进化革命,因为我们将看到它与众不同之处,因为它在所有物种中都有着共同的模式,并且沿着不同的途径在无脊椎动物中受到赞赏的沿不同途径的持续改进过程不在这里。与无脊椎动物不同,它来自神经外胚层。
脊椎动物的眼睛没有显示出在大脑,耳朵或心脏中看到的不断增加的分化,并且鱼眼本质上与鸟类或哺乳动物的眼睛一样复杂且完全发育。这只是一个谜团,尽管对我们来说最有趣,但它仍然是一个谜,即对进化机制的理解继续从没有骨架的动物扔到拥有如此完整骨架的其他人手中。
从原理上讲,根据栖息地的不同,脊椎动物的眼睛分为三种类型:
-仅在幼体阶段才适应水生环境的鱼鳞,鱼类和两栖动物。
-蜥脚类动物,爬行动物和鸟类,适应空气环境。
-哺乳动物的,矛盾的。
视网膜的基本元素在所有脊椎动物中都具有相同的结构,尽管视光而定,视杆和视锥之间的关系也有所不同。有人认为视锥细胞是原始的视觉细胞,视杆是后来的元素,这是由于视紫红质与发育相关的trans变所致,这种trans变首先发生在深海鱼类中。在其余的眼部结构中,脊椎动物存在着很大的差异,这是由于它们适应了非常多样化的环境:最初的眼睛适应了浅水区,但后来视情况而演化为存在于深海中。在河流中,沼泽中的泥土中,地面和空气中以及夜晚,昏暗或半暗的洞穴或最强烈的白天。当必须对运动进行感知时,它还可以实现全景视觉;对于距离的提取计算,它可以实现立体视觉,可以在简单的模糊感知和最精确的分辨率或最复杂的色彩视觉之间进行振荡。
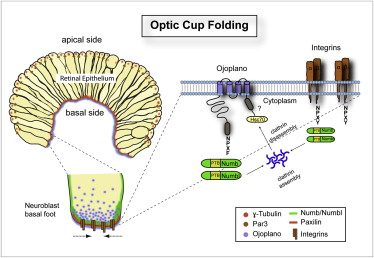
图31,光学球罩的折叠控制
忽略少数气旋,我们就可以总结出鱼,两栖动物,爬行动物,鸟类和哺乳动物的眼睛的主要特征。
至。鱼。
角膜实际上没有屈光度值,因此眼球往往非常大且具有折射晶体。角膜变平以更好地抵抗水压。水深越深,眼球的大小就越大,以至于在无光的深渊空间中,眼睛退化并几乎消失。甚至在鳗鱼这样的物种中,随着动物从欧洲或美洲时代到大西洋的猿猴海旅行,它们的眼睛也会长大,在那里它们繁殖并死亡。非常富含杆的视网膜通常是无血管的。眼睛运动受到很大限制,最好通过头部和全身运动来实现固定。我们已经知道视觉色素的奇异之处,即淡水鱼是由维生素A2制成的,并且像鲑鱼一样,也从海洋迁移到河流,从而改变了视觉色素。
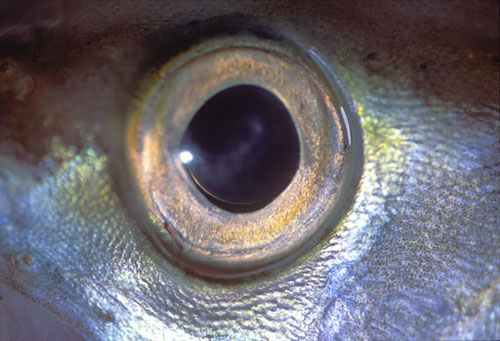
图32,鱼眼
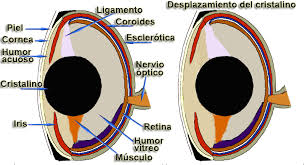
图33,鱼眼示意图
b。两栖动物
从水生环境到空气环境的通道使角膜具有屈光功能,变得很凸。晶状体比鱼小,可以通过前后移位来适应。出现保护性结构,例如眼睑,腺体和泪管。

图34,普通蛙眼
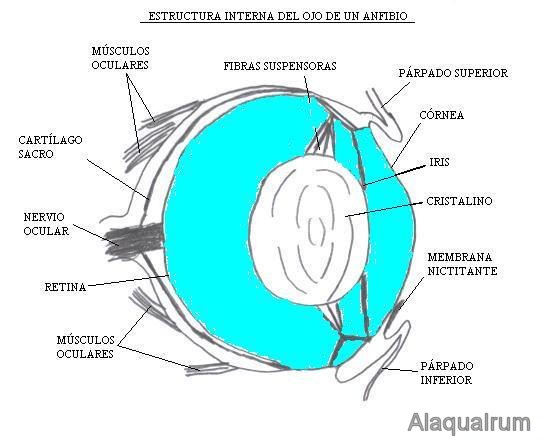
图35,两栖动物的眼睛图
C。爬虫类
调节是通过晶状体的变形来进行的,该变形需要存在附着在角膜周围的睫状肌。视网膜是无血管的,并以脉络膜或视网膜外的血管膜的营养为代价。他们有第三只眼皮。
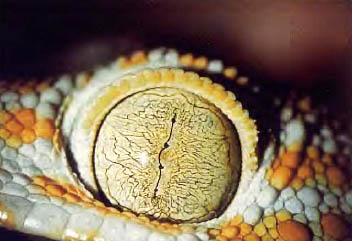
图36,爬行动物的眼睛
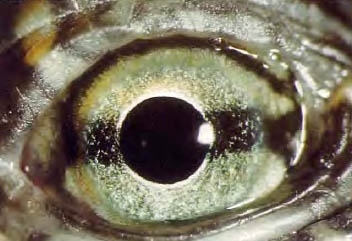
图37,海龟的眼睛
d。鸟类。
眼睛通常比因晶状体变形而具有调节机制的眼睛大。视网膜很厚,根据生活习惯,有大量的感光细胞,视锥细胞和/或视杆较丰富。视网膜由果胶(一种对应于视盘的细长血管膜)滋养。在某些类型中,有两个中央凹,一个用于远视,另一个用于近视。
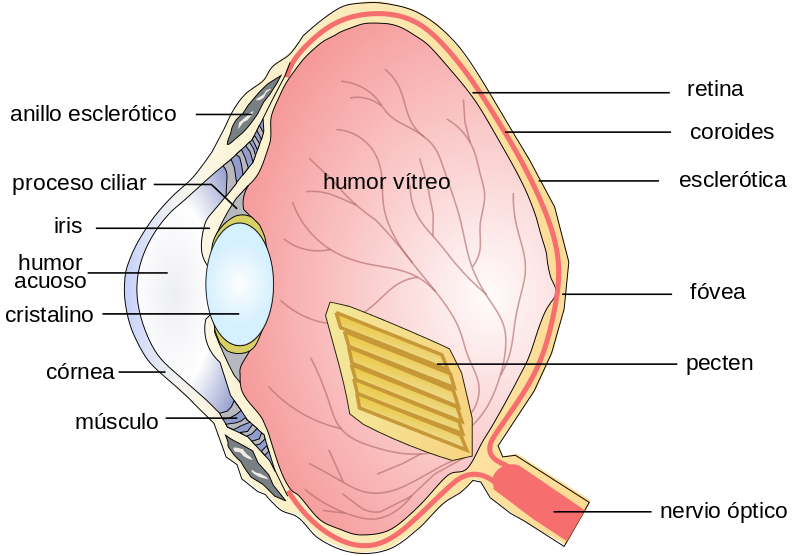
图38,鸟眼的解剖图

图39,猫头鹰眼
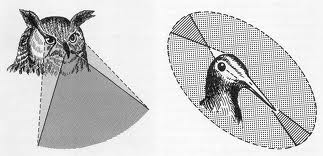
图40,鸟类视野
和。哺乳动物
鸭嘴兽的单眼与爬行动物非常相似,有袋类动物的视线介于爬行动物和胎盘之间。这是种类最多,种类最多的子类,包括十五个科:食虫类,手翅类,皮肤癣菌,灵长类动物(具有多个子类和科),啮齿动物,食肉动物,关节突动物,过鞭毛基动物,象鼻动物,警报器,鲸类...
胎盘的眼睛在三个基本方面与其余脊椎动物不同:
-透明状动脉的发育和随后的向内卷入,在许多情况下会导致视网膜内血管形成。
-虹膜的中胚层的外观。
-由于晶状体囊的变形而在容纳机构中。
自然地,栖息地和生活习惯在其余的眼部结构中产生微小的变化:在水生环境中,眼球结构几乎呈球形结晶,脉络膜和视网膜之间存在或不存在绒毡层,瞳孔和括约肌形状的巩膜厚度(在鲸鱼中为眼径的3/4),在视网膜的血管化过程中(全部,部分或不存在:全血管性,中血管性和血管性视网膜)在视网膜棒和视锥细胞的比例。

图41,鲸鱼眼
最有趣的方面之一(因为对每个眼部结构的每个顺序,子顺序和族的详细描述将使我们走得太远)是光纤的讨论。
在其他脊椎动物中,几乎全部消散,这使得双眼视野可以跨越360°,但将立体视觉的局限性限制为不如哺乳动物。在这些动物中,尤其是在胎盘中,他开始看到与眼睛额叶有关的直接纤维。马的比例为马的1/6,猫的比例为1/4至1/3,高等灵长类的比例为1/3,人的比例为1/2。这个过程意味着失去了双眼视野的扩大,但是获得了精确的立体视觉。据推测,这对于生活在树栖环境中的动物来说是必要的,在这种环境中,分支的运动需要对距离进行适当的计算,并与手成为有力器官有关。
图42,讨论人类的视觉通路
我们已经指出,只有在哺乳动物中,大脑皮层的区域才会出现视觉冲动到达并被分析的区域。即使这样,除了人以外,该位置仍然不精确,在大多数哺乳动物中,视皮层的消融不会引起完全失明。然而,在人类中,唯一保持皮层下的视觉活动是瞳孔。
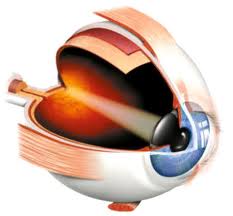
图43,人眼示意图
视觉功能
环境和生活习惯(最好是白天或晚上的活动)会在视觉功能的各个方面产生广泛的变化:光线感知,色彩感,形式感,立体视觉...
一个例子总结了在脊椎动物中实现调节的不同机制:
静态机制:
针孔瞳孔。
复制的光学系统。
插入硝化膜。
复制的视网膜。
倾斜的视网膜。
粗糙的视网膜。
接收元件的长度很大。
动态机制:
由角膜压力或自身肌肉组织引起的晶状体向前或向后移位。
晶状体变形,这在哺乳动物中最终归因于囊的弹性及其不同的厚度。
鸟类的视敏度达到10弧度的分辨率,是人类的三倍。
单眼视场的范围从变色龙的30°到马的215°。在淹没的鱼类中,水生和空中视觉相结合,但是受到水面总折射的限制。正如我们之前所提到的,双目望远镜在动物身上可以覆盖360°,可以捕食,在掠食者中可以减少。在眼睛侧面定位的鱼中,考虑到球形表面,盲区可能很小。
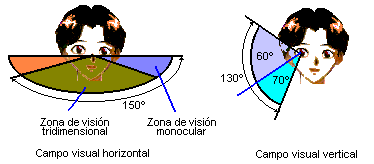
图44,人的视野
同时,由光轴和中线形成的角度变化很大,例如在哺乳动物中仅共轭眼运动的极限。
5.数字索引。
图1,植物的光与新陈代谢。
图2,女性性周期。
图3,向日葵的植物趋光性。
图4,向光性机理(生长素)。
图5,变形虫在阿米巴中的运动。
图6,飞蛾的趋光性。
图7,在裸藻中的气候轴。
图8,蠕虫中的对光轴。
图9,复眼。
图10,鸟的menotaxis。
图11,蜜蜂的驱虫作用。
图12,唯一,色素沉着的保护功能。
图13,孔雀的色素沉着功能。
图14,棕褐色(黑色素)。
图15,变色龙(guanophores)。
图16,猩猩的手眼协调能力。
图17,人的视觉通路:视觉皮层化。
图18,感光体和颜料的演变。
图19,气管酮方案。
图20,气管单胞菌。
图21,紫丁香。
图22,Stilaria L.
图23,平眼,紫丁香。
图24,骨菌。
图25,eye骨SP杯中的眼睛。
图26,螺旋果皮。
图27,螺旋P中的囊泡眼
图28,复眼。
图29,复眼方案。
图30,孔眼的结构。
图31,光学球罩的折叠控制。
图32,鱼眼。
图33,鱼眼示意图。
图34,普通的青蛙眼。
图35,两栖动物的眼睛图。
图36,爬行动物的眼睛。
图37,海龟的眼睛。
图38,鸟眼的解剖图。
图39,猫头鹰眼。
图40,鸟类视野。
图41,鲸鱼眼。
图42,讨论了人类的视觉通路。
图43,人眼,示意图。
图44,人的视野。
6.参考书目。
- 进化中的眼睛,《眼科学系统》第一卷,斯图尔特·杜克·埃尔德爵士,C.V。莫斯比公司(Mosby Company),1958年。
- 动物之眼》,迈克尔·F·兰德,丹·埃里克·尼尔森,第二版,2012年,牛津动物生物学系列。
- Webvision,视网膜和视觉系统的组织,http://webvision.med.utah.edu/
- Shaun P Collin和Ann EO Trezise的《脊椎动物彩色视觉的起源》,《临床和实验验光》,2004年。
- 眼睛进化的“劳动分工”模型,Detlev Arendt,Harald Hausen和GünterPurschke,Phil。反式R.Soc.B 2009 364,doi:10.1098 / rstb.2009.0104,2009年8月31日发布。
- 早期脊椎动物感光细胞的进化,Shaun P. Collin,Wayne L. Davies,Nathan S. Hart和David M. Hunt,Phil。反式R.Soc.B 2009 364,doi:10.1098 / rstb.2009.0099,2009年8月31日发布。
- 鸟类和哺乳动物中视觉色素的演变和光谱调整,David M. Hunt,Livia S. Carvalho,Jill A. Cowing和Wayne L. Davies,Phil。反式R.Soc.B 2009 364,doi:10.1098 / rstb.2009.0044,2009年8月31日发布。
- 哺乳动物颜色视觉的演变,Gerald H. Jacobs,菲尔。反式R.Soc.B 2009 364,doi:10.1098 / rstb.2009.0039,2009年8月31日发布。
- 趋光性的演变,GáspárJékely,菲尔。反式R.Soc.B 2009 364,doi:10.1098 / rstb.2009.0072,2009年8月31日发布。
- 脊椎动物视觉类维生素A循环的进化和起源,Takehiro G. Kusakabe,Noriko Takimoto,Minghao Jin和Motoyuki Tsuda,Phil。反式R.Soc.B 2009 364,doi:10.1098 / rstb.2009.0043,2009年8月31日发布。
- 光电导和眼睛的演化,Trevor D. Lamb,Detlev Arendt和Shaun P. Collin,菲尔。反式R.Soc.B 2009 364,doi:10.1098 / rstb.2009.0106,2009年8月31日发布。
- 脊椎动物视网膜光感受态的进化,Trevor D. Lamb,菲尔。反式R.Soc.B 2009 364,doi:10.1098 / rstb.2009.0102,2009年8月31日发布。
- 眼睛和视觉引导行为的演变,Dan-Eric Nilsson,菲尔。反式R.Soc.B 2009 364,doi:10.1098 / rstb.2009.0083,2009年8月31日发布。
- 辐照度检测的演变:黑视蛋白和非视觉视蛋白,Stuart N. Peirson,Stephanie Halford和Russell G. Foster,Phil。反式R.Soc.B 2009 364,doi:10.1098 / rstb.2009.0050,2009年8月31日发布。
- 视蛋白和光转导的演变,吉田义德和松山健,Phil。反式R.Soc.B 2009 364,doi:10.1098 / rstb.2009.0051,2009年8月31日发布。