AUTHOR: MANUEL D. VALDEARENAS MARTIN
The visual system is the specific receptor for light stimuli, which not only act on animals: let us recall photosynthesis in plants, mediated by chlorophyll, which transforms carbon dioxide and water into carbohydrates and oxygen, with accumulation of energy.
Light energy can act on living organisms in four different ways: influencing metabolic processes, affecting movements, being related to pigmentation or giving rise to the mechanisms of vision itself (physical, chemical, neural or psychological).
1. EFECTOS DE LA LUZ SOBRE EL METABOLISMO.
The daily rhythm of night and day and the seasonal variations of this rhythm cause the photo periodicity that occurs throughout the animal scale and that influences even the sexual cycle.
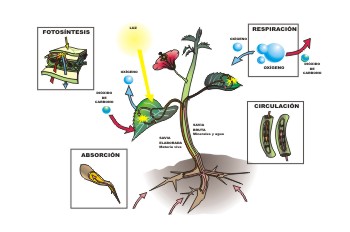
Figure 1, light and plant metabolism
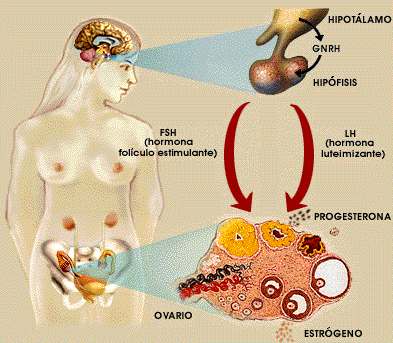
Figure 2, Female Sexual Cycle
2. EFFECTS OF LIGHT ON MOVEMENT.
They are of three different types:
- PHOTOTROPISM
Phototropism is more typical of plants and consists of the movement towards the light or away from it of the sessile organs (positive, negative or transverse heliotropism).

Figure 3, plant phototropism in sunflowers
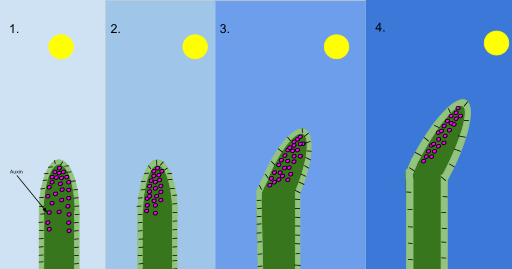
Figure 4, phototropism mechanism (auxin)
- PHOTOQUINESIS
Alteration of motility without directional orientation. A distinction is made between orthokinesis (acceleration or slowing down of movements) and clinokinesis (deviation of movements towards or away from light).
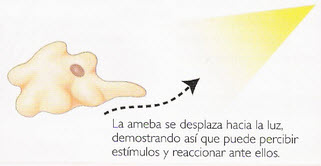
Figure 5, clinokinesis in amoeba
- PHOTOTAXIS
Alteration of motility with directional orientation. It requires receivers that can distinguish the direction of the light source.

Figure 6, phototaxis in moths
The phototaxis is divided into:
Clinotaxis
It requires the existence of a receptor organ that compares the intensity of successive stimuli to direct the displacement in relation to the light in an irregular way (for example the orientation of the flagellates).
Figure 7, clinotaxis in euglena
Tropotaxis
It requires two symmetric receptors and the movement, more exact, is made by comparing the intensity of the stimulation of each of them (for example, the displacement of the worms).
Figure 8, tropotaxis in worm
Telotaxis
Orientation is carried out directly towards or from the light source. Although a single receptor is sufficient, it must be complex and composed of several elements. In addition, it is necessary to have a central nervous organization that analyzes stimuli and inhibits unnecessary or less intense ones (compound eye of insects).
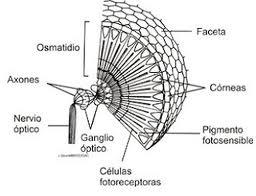
Figure 9, compound eye
Menotaxis
It includes the ability to move not only to or from the light, but also at an angle (light compass reaction). The navigation system of birds and the possible orientation of polarized second light (dance of the bees) are based on it. It also includes orientation in front of an image and the dorsal or ventral light reaction that helps air or aquatic navigation.
Figure 10, menotaxis in birds
Mnemotaxis
That includes the existence of a visual memory with which current stimuli are compared.
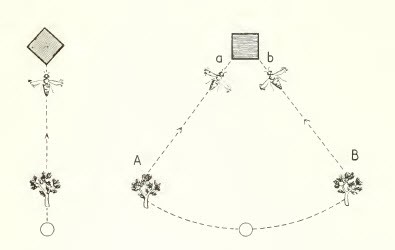
Figure 11, mnemotaxis in bees
3. EFFECTS OF LIGHT ON PIGMENTATION.
The pigment that exists to absorb light or to filter it and has a less frequent protective (cryptic) or demonstrative (faneric) function and of social or reproductive value.

Figure 12, sole, Protective function of pigmentation

Figure 13, Peacock, phaneric function of pigmentation
It requires the existence of organized cells or cell groups: chromatophores. The pigment can be melanin (melanophores), it will be composed of fat-soluble carotenoids (lipophors) or guanine (guanophores), the last two being of a great variety of colors and iridescent.

Figura 14, sepia (melanóforos)

Figure 15, Chameleon (guanophores)
The action of light on the pigment can be of three types:
Direct on the melanophores of the skin or even on retinal or iris ectoderm: direct pupillary contraction in fish and amphibian cephalopods.
Hint through the eye.
Indirect through the midbrain (some fish) or the pineal gland (lamprey, teleost fish).
4. VISION PROPERLY SAID.
The essential and primitive function of vision is the control of movements to achieve as easily as possible an optimal environment. This is used to avoid obstacles, to pursue prey or flee from predators and in man requires a close relationship between the eyes and the vestibular apparatus..

Figure 16, hand-eye coordination in orangutan
Higher levels of visual function are, by their very nature, conscious, although in lower animals (eg, the ant's gait) they are not, and neither naturally in machines regulated by cybernetic feedback mechanisms. The process of “awareness” of vision has coincided, or has been due, to the migration of the terminal station from the basal nuclei to the cortex (which is a recent fact in the history of evolution).
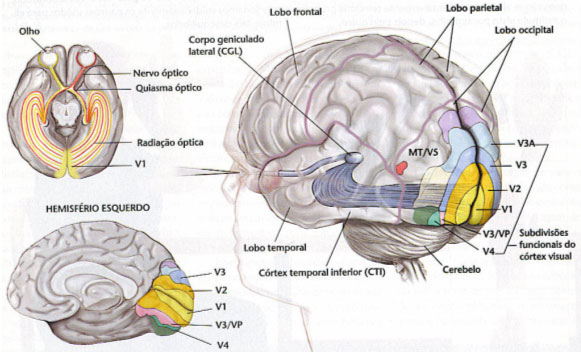
Figure 17, optic pathways in man: corticalization of vision
THE EVOLUTION OF THE VISUAL APPARATUS
Although in lower animals there is diffuse light sensitivity throughout the ectoderm, soon part of it specializes in light reception, which is limited to photoreceptors. These are characterized by the presence of pigments: melanin, which is chemically inert and acts only as an absorbent, and the visual pigments themselves. All these are based on carotenoids associated or not with proteins, which are also found, in addition to chlorophyll, in the plant kingdom (xanthophyll). When the level of the eye with optical imaging capacity is reached, all pigments are specifically related to vitamin A (A1 or A2).
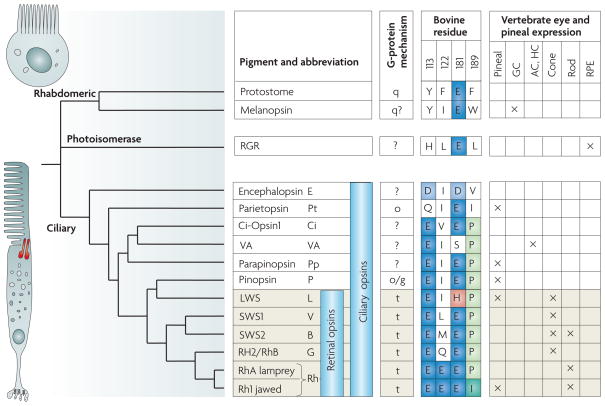
Figure 18, evolution of photoreceptors and pigments
In unicellular organisms, diffuse light sensitivity can be concentrated in a point that is associated with cilia or flagella (motor function). In flagellates this "eye point" or "stigma" has pigment and occasionally a retractable structure that concentrates light and acts, like a lens (crystalline).
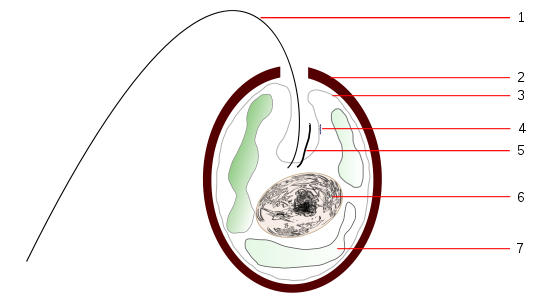
Figure 19, scheme of tracheloomone
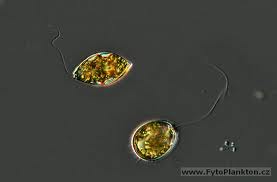
Figure 20, trachelomonas
In multicellular beings, some ectodermal cells soon specialize in light reception as bipolar cells (a distal receiving end and a proximal transmitting end) or apolar cells of ellipsoidal shape. From here begins the evolution of the eye in invertebrates.
THE EYE OF THE INVERTEBRATES
It derives from the surface ectoderm and secondarily connects with the nervous system.
It can be simple, either unicellular or multicellular, or compound.
- Simple eye an ocellus.
It consists of one or more light-sensitive cells that act without a functional association.
When it is multicellular it can be still subepithelial (just a collection of photosensitive cells) or in a more advanced stage invaginate epithelial. This last type passes through the flat, cupular (formation of a dark chamber) and finally vesicular phases, which is still perfected in the eye of the cephalopods in which a thickening of the epithelium already forms a “shadow” of the lens, and there is also a contractile pupil.

Figure 21, stilaria lacustris
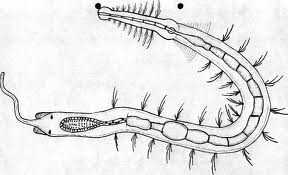
FFigure 22, Anatomia Stilaria L
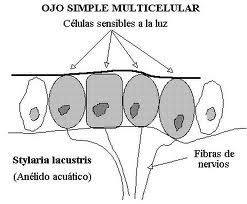
Figure 23, flat eye, stilaria L.

Figure 24, patella sp.
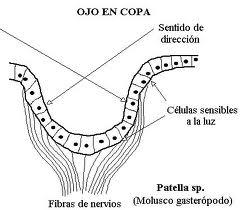
Figure 25, eye in cup in patella SP

Figure 26, helix pomatia
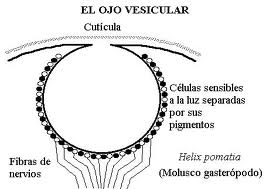
Figure 27, vesicular eye in helix P.
Although the existence of an inverted retina (in which the photoreceptor elements are in the deepest layer and the transmitters on the surface, which allows a better nutrition of the former) is characteristic of vertebrates, some mollusks and arachnids already have this new retinal organization, instead of the "poured" or direct retina. This change in the orientation of the retina is one of the most difficult processes to explain in light of the theory of continuous evolution. In addition, in animals that have to live in poor lighting conditions, a reflective crystalline layer or tapetum appears behind the retina..
- Compound eye.
In the compound eye the sensory elements are structurally and sensory grouped. Each one of them, called ommatidium, consists of a corneal facet, a crystalline cone, an iridian pigmented cell and a retinae composed of photoreceptors that end in a prolongation common to several of them, called a rhabdom, in which transmission begins. of impulses to nerve centers.
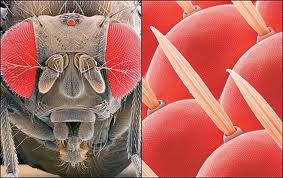
Figure 28, compound eye
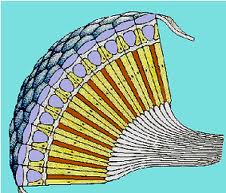
Figure 29, compound eye scheme
The compound eye, peculiar to arthropods, adopts different varieties in arachnids, crustaceans and insects. Although it only allows to achieve a reduced visual acuity, variable depending on whether there is a greater or lesser concentration of the retina on the rhabdomes, the compound eye has the ability to analyze the plane of polarization of light, an essential function for orientation in space.
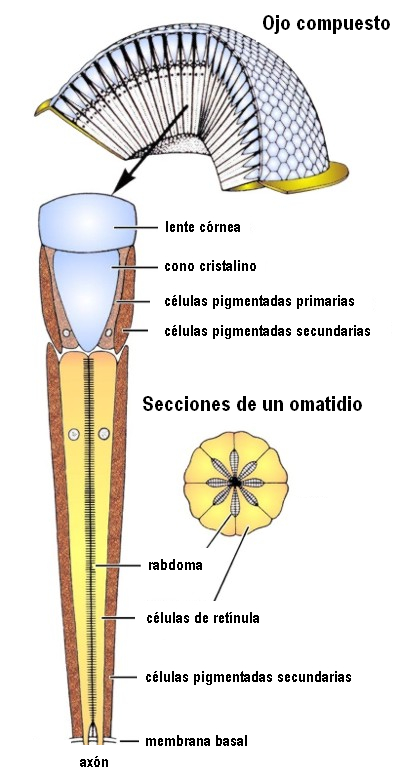
Figure 30, structure of an ommatidium
THE EYE OF THE VERTEBRATES
It supposes an evolutionary revolution since, with the differences that we will see, it has a common pattern in all species and the continuous process of improvement, along different routes, that is appreciated in invertebrates is not assisted here. Unlike invertebrates it derives from the neural ectoderm.
The vertebrate eye does not show the increasing differentiation seen in the brain, ear or heart, and the eye of a fish is essentially as complex and fully developed as that of a bird or mammal. This is just one, although for us the most interesting, of the mysteries that the understanding of evolutionary mechanisms continues to throw from animals without a skeleton to others that possess it so complete.
Schematically the eye of vertebrates is differentiated into three types depending on the habitat:
- That of ichthyosids, fish and amphibians, adapted to an aquatic environment, in the latter only in the larval phase.
- That of the Sauropsids, reptiles and birds, adapted to an air environment.
- The one with the mammals, ambivalent.
The fundamental element, the retina, has the same structure in all vertebrates, although the relationship between rods and cones varies depending on the lighting. Some think that the cones are the primitive visual cell and that the rods are a later element due to a developmentally related transmutation of rhodopsin, which occurred first in deep-sea fish. In the rest of the ocular structures, there are considerable differences in vertebrates that are the consequence of adaptation to very diverse environments: initially the eye adapted to shallow waters but later it has evolved, depending on the case, to existence in abysmal waters. in rivers, in the mud of swamps, on the ground and in the air as well as at night, twilight or semi-darkness of caves or the most intense daylight. It has also achieved a panoramic vision when the perception of movement is essential or a stereoscopic vision when it is an extract calculation of distances, oscillating between a simple vague apperception and the most exact resolution capacity or the most complex chromatic vision..
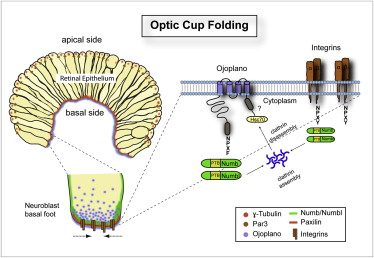
Figure 31, control of the folding of the optical dome
Disregarding the few cyclostomes, we can thus summarize the main characteristics of the eye of fish, amphibians, reptiles, birds and mammals..
a. Fishes.
La córnea carece prácticamente de valor dióptrico, de modo que el globo ocular tiende a ser muy grande y cristalino refringente. La córnea es aplanada para resistir mejor la presión del agua. El tamaño del globo ocular es mayor cuanto mayor lo es la profundidad de las aguas, de tal forma que en los espacios abisales, desprovistos de luz, el ojo degenera y casi desaparece. Incluso en especies como la anguila el ojo crece al desplazarse el animal desde los días de Europa o América hasta el mar de los sargazos en el atlántico donde procrean y mueren. La retina muy rica en bastones suele ser avascular. Los movimientos oculares son muy restringidos y la fijación se consigue preferentemente con movimientos de todo cuerpo y la cabeza. Ya conocemos la curiosa diferencia en los pigmentos visuales, que los peces de agua dulce están constituidos a base de vitamina A2 y en los que, como los salmones, emigran de los mares a los ríos y cambia consiguientemente el pigmento visual.
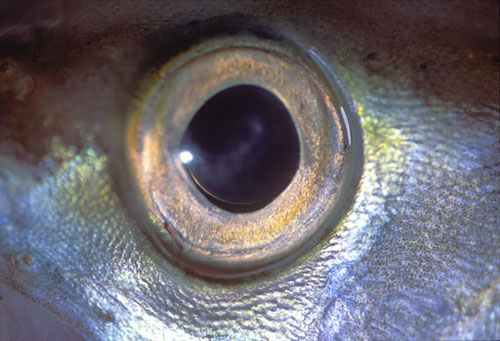
Figure 32, fisheye
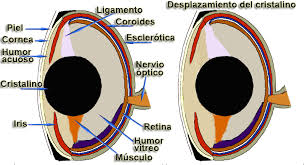
Figure 33, fish eye schematic
b. Amphibians.
The passage from the aquatic to the air environment allows the cornea to adopt dioptric functions, becoming strongly convex. The lens is smaller than the fish, allows accommodation through anteroposterior displacements. Protective structures such as the eyelids, glands and tear ducts appear.

Figure 34, common frog eye
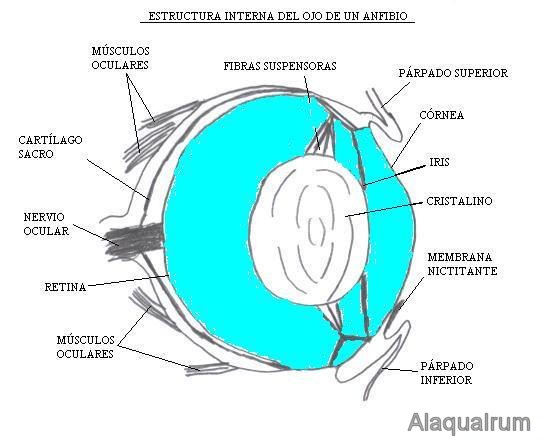
Figure 35, diagram of the eye of an amphibian
c. Reptiles.
The accommodation is carried out through a deformation of the lens, which requires the existence of a ciliary muscle that is attached to the periphery of the cornea. The retina is avascular and is nourished at the expense of the choroid or an extraretinal avascular membrane. They have a third eyelid.
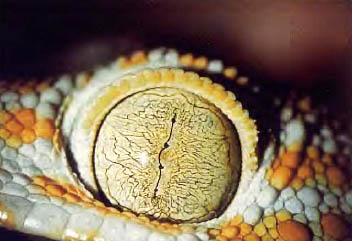
Figure 36, reptile eye
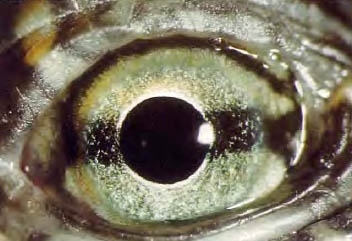
Figure 37, sea turtle eye
d. Birds.
The eye is usually larger than the one that appears to have an accommodative mechanism through deformation of the lens. The retina is thick, with a great abundance of photoreceptors, cones and rods, more abundant according to life habits. The retina is nourished by a Pecten, an elongated vascular membrane that corresponds to the optic disc. In some types there are two foveas, one for distant vision and one for near vision.
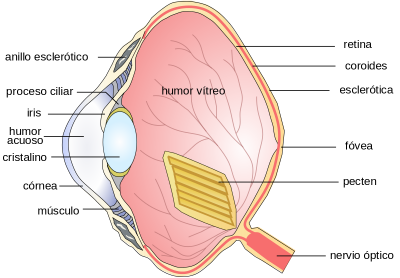
Figure 38, anatomical scheme of the eye of birds

Figure 39, owl eye
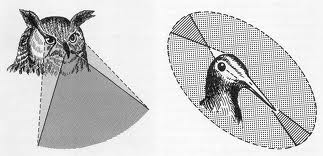
Figure 40, visual field in birds
e. Mammals.
The eye of monotremes, platypus, is very similar to that of reptiles, that of marsupials has an intermediate structure between that of reptiles and that of placentates. This is the most abundant and varied subclass and comprises fifteen orders: insectivores, chiropterans, dermatoptera, primates (with several suborders and families), rodents, carnivores, artiodactyls, perissodactyls, proboscids, sirens, cetaceans…
The eye of the placentates differs from the rest of the vertebrates in three fundamental aspects:
- The development and subsequent involution of the hyaloid artery, which in many cases leads to intraretinal vascularization.
- In the appearance of the mesodermal layer of the iris.
- In the accommodative mechanism due to a deformation of the lens capsule.
Naturally, habitat and life habits impose minor variations in the rest of the ocular structures: almost spherical crystalline in those that live in an aquatic environment, presence or absence of a tapetum located between the choroid and the retina, variations in the thickness of the sclera (which is 3/4 of the ocular diameter in the whale), the shape of the pupil and the sphincter, the vascularization of the retina (total, partial or absent: holoangiotic, mesoangiotic and anangiotic retinas) and in the ratio of retinal rods and cones.
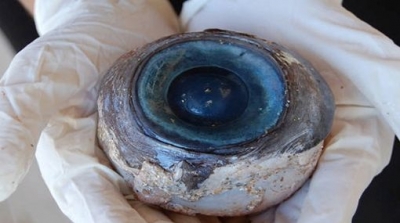
Figure 41, whale eye
One of the most interesting aspects (since the detailed description of each ocular structure in each order, suborder and family would take us too far) is that of the decussation of the optical fibers.
In the other vertebrates, decussation is practically total, which allows a binocular visual field that can span 360 ° but limits stereoscopic vision to less exact mechanisms than mammals. In these, especially in the placentates, he begins to see direct fibers in relation to the frontalization of the eyes. The proportion is 1/6 of the total in the horse, 1/4 to 1/3 in the dog and cat, 1/3 in the higher primates and almost 1/2 in the man. This process implies a loss of the extension of the binocular visual field but the acquisition of an exact stereoscopic vision. It has been speculated that this began to be necessary in animals that lived in an arboreal environment, in which the movements of the branch require an adequate calculation of the distances and was associated with the development of the hand as a prehensile organ.
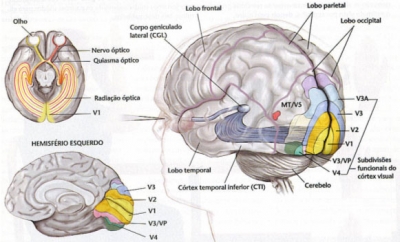
Figure 42, decussation of the visual pathways in man
We have already pointed out that only in mammals does an area of the cerebral cortex appear where visual impulses arrive and are analyzed. Even so, except in man, this location is still imprecise and in most mammals ablation of the visual cortex does not cause complete blindness. In man, however, the only visual activity that remains subcortical is the pupillary.
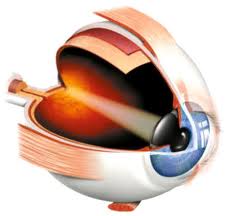
Figure 43, human eye, scheme
VISUAL FUNCTION
The environment and life habits, preferably daytime or nighttime activity, impose a wide range of variations in all aspects of visual function: light perception, color sense, sense of form, stereoscopic vision…
An example is a summary of the different mechanisms by which accommodation is achieved in vertebrates:
Static mechanisms:
A pinhole pupil.
A duplicated optical system.
The interposition of a nictitating membrane.
A duplicated retina.
A ramping retina.
A rough retina.
A great length of the receiving elements.
Dynamic mechanisms:
Displacement of the lens forwards or backwards, induced by corneal pressure or by own musculature.
Lens deformation, which in mammals is ultimately due to the elasticity of the capsule and its different thickness.
La agudeza visual llega en las aves alcanzar una resolución de 10” de arco, tres veces superior a la del hombre.
The monocular field of view ranges from 30 ° for the chameleon to 215 ° for the horse. In submerged fish, aquatic and aerial vision are combined, but subject to the restriction imposed by the total refraction of the water surface. The binocular, as we mentioned before, covers 360 ° in animals that can be prey and is reduced in predators. In fish with laterally positioned eyes, the blind area, considering a spherical surface, can be very small.
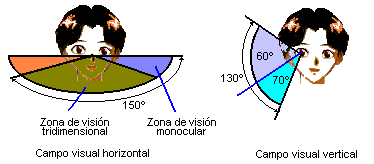
Figure 44, visual field in man
At the same time there is a great variety with the angle formed by the optic axes and the midline, as in the limit of only conjugated ocular motility in mammals.
5. ÍNDICE DE FIGURAS.
Figure 1, light and metabolism of plants.
Figure 2, Female sexual cycle.
Figure 3, plant phototropism in sunflowers.
Figure 4, phototropism mechanism (auxin).
Figure 5, clinokinesis in amoeba.
Figure 6, phototaxis in moths.
Figure 7, clinotaxis in euglena.
Figure 8, tropotaxis in worm.
Figure 9, compound eye.
Figure 10, menotaxis in birds.
Figure 11, mnemotaxis in bees.
Figure 12, sole, Protective function of pigmentation.
Figure 13, Peacock, phaneric function of pigmentation.
Figure 14, sepia (melanophores).
Figure 15, Chameleon (guanophores).
Figure 16, hand-eye coordination in orangutan.
Figure 17, optic pathways in man: corticalization of vision.
Figure 18, evolution of photoreceptors and pigments.
Figure 19, scheme of trachelomone.
Figure 20, trachelomonas.
Figure 21, stilaria lacustris.
Figure 22, Stilaria L. anatomy
Figure 23, flat eye, stilaria L.
Figure 24, patella sp.
Figure 25, eye in cup in patella SP.
Figure 26, helix pomatia.
Figure 27, vesicular eye in helix P.
Figure 28, compound eye.
Figure 29, compound eye scheme.
Figure 30, structure of an ommatidium.
Figure 31, control of the folding of the optical dome.
Figure 32, fisheye.
Figure 33, schematic of the fish eye.
Figure 34, common frog eye.
Figure 35, diagram of the eye of an amphibian.
Figure 36, reptile eye.
Figure 37, sea turtle eye.
Figure 38, anatomical diagram of the eye of birds.
Figure 39, owl eye.
Figure 40, visual field in birds.
Figure 41, whale eye.
Figure 42, decussation of the visual pathways in man.
Figure 43, human eye, schematic.
Figure 44, visual field in man.
6. BIBLIOGRAPHY.
- The eye in evolution, System of Ophthalmology Volumen I, Sir Stewart Duke-Elder, The C.V. Mosby Company, 1958.
- Animal Eyes, Michael F. Land, Dan-Eric Nilsson, 2ª Edition, 2012, Oxford Animal Biology Series.
- Webvision, The Organization of the Retina and Visual System, http://webvision.med.utah.edu/
- The origen of colour visión in vertebrates, Shaun P Collin and Ann EO Trezise, Clinical and experimental optometry, 2004.
- The 'division of labour' model of eye evolution, Detlev Arendt, Harald Hausen and Günter Purschke, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0104, published 31 August 2009.
- The evolution of early vertebrate photoreceptors, Shaun P. Collin, Wayne L. Davies, Nathan S. Hart and David M. Hunt, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0099, published 31 August 2009.
- Evolution and spectral tuning of visual pigments in birds and mammals, David M. Hunt, Livia S. Carvalho, Jill A. Cowing and Wayne L. Davies, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0044, published 31 August 2009.
- Evolution of colour vision in mammals, Gerald H. Jacobs, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0039, published 31 August 2009.
- Evolution of phototaxis, Gáspár Jékely, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0072, published 31 August 2009.
- Evolution and the origin of the visual retinoid cycle in vertebrates, Takehiro G. Kusakabe, Noriko Takimoto, Minghao Jin and Motoyuki Tsuda, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0043, published 31 August 2009.
- The evolution of phototransduction and eyes, Trevor D. Lamb, Detlev Arendt and Shaun P. Collin, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0106, published 31 August 2009.
- Evolution of vertebrate retinal photoreception, Trevor D. Lamb, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0102, published 31 August 2009.
- The evolution of eyes and visually guided behaviour, Dan-Eric Nilsson, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0083, published 31 August 2009.
- The evolution of irradiance detection: melanopsin and the non-visual opsins, Stuart N. Peirson, Stephanie Halford and Russell G. Foster, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0050, published 31 August 2009.
- Evolution of opsins and phototransduction, Yoshinori Shichida and Take Matsuyama, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0051, published 31 August 2009.