AUTOR: MANUEL D. VALDEARENAS MARTIN
El sistema visual es el receptor específico de los estímulos luminosos, que no sólo actúan sobre los animales: recordemos en los vegetales la fotosíntesis, mediada por la clorofila, que transforma el dióxido de carbono y el agua en hidratos de carbono y oxígeno, con acumulación de energía.
La energía luminosa puede actuar sobre los organismos vivos de cuatro formas distintas: influenciando los procesos metabólicos, afectando los movimientos, relacionándose con la pigmentación o dando lugar a los mecanismos de la visión propiamente dicha (físicos, químicos, neurales o psicológicos).
1. EFECTOS DE LA LUZ SOBRE EL METABOLISMO.
El ritmo diario de noche y día y las variaciones estacionales de este ritmo ocasiona la foto periodicidad que se presenta en toda la escala animal y que influencia hasta el ciclo sexual.
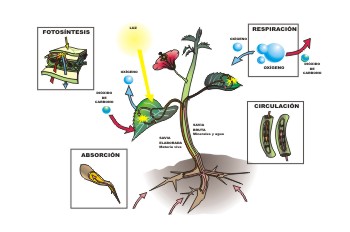
Figura 1, luz y metabolismo de las plantas
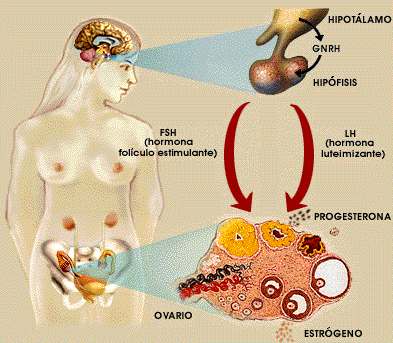
Figura 2, Ciclo sexual femenino
2. EFECTOS DE LA LUZ SOBRE EL MOVIMIENTO.
Son de tres tipos diferentes:
- FOTOTROPISMO
El fototropismo es más propio de los vegetales y consiste en el desplazamiento hacia la luz o alejamiento de ella de los órganos sésiles (heliotropismo positivo, negativo o transverso).

Figura 3, fototropismo vegetal en girasoles
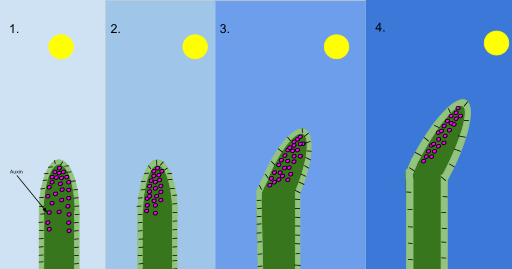
Figura 4, mecanismo fototropismo (auxina)
- FOTOQUINESIS
Alteración de la motilidad sin orientación direccional. Se distingue entre la ortoquinesis (aceleración o enlentecimiento de los movimientos) y la clinoquinesis (desviación de los movimientos hacía o lejos de la luz).
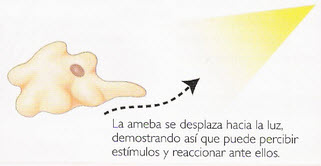
Figura 5, clinoquinesis en la ameba
- FOTOTAXIS
Alteración de la motilidad con orientación direccional. Precisa receptores que puedan distinguir la dirección de la fuente luminosa.

Figura 6, fototaxis en polillas
La fototaxis se divide en:
Clinotaxis
Requiere la existencia de un órgano receptor que compara la intensidad de estímulos sucesivos para dirigir el desplazamiento en relación a la luz de una manera irregular (por ejemplo la orientación de los flagelados).
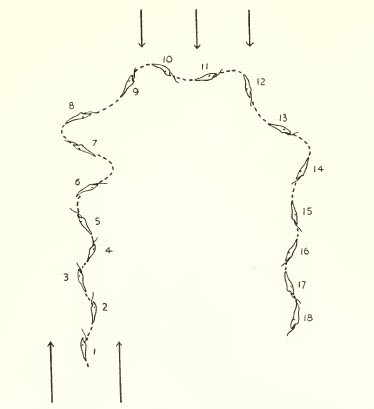
Figura 7, clinotaxis en euglena
Tropotaxis
Requiere dos receptores simétricos y el movimiento, más exacto, se realiza al comparar la intensidad de la estimulación de cada uno de ellos (por ejemplo el desplazamiento de los gusanos).
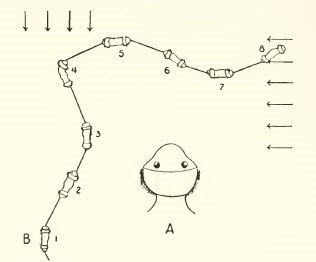
Figura 8, tropotaxis en gusano
Telotaxis
La orientación se efectúa directamente hacia o desde la fuente luminosa. Aunque basta un solo receptor, éste debe ser complejo y compuesto de varios elementos. Además hay que contar con una organización nerviosa central que analice los estímulos e inhiba los innecesarios o menos intensos (ojo compuesto de los insectos).
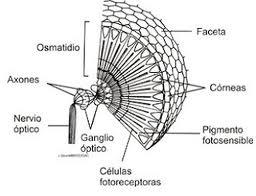
Figura 9, ojo compuesto
Menotaxis
Incluye la capacidad de desplazarse no sólo desde o hacia la luz, sino también de forma angular (reacción del compás luminoso). En ella se basa el sistema de navegación de las aves y la posible orientación segunda luz polarizada (baile de las abejas). Incluye también la orientación frente a una imagen y la reacción luminosa dorsal o ventral que ayuda a la navegación aérea o acuática.
Figura 10, menotaxis en aves
Mnemotaxis
Que incluye la existencia de una memoria visual con la que se compara los estímulos actuales.
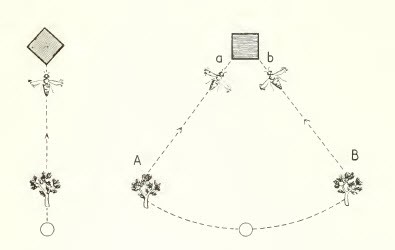
Figura 11, mnemotaxis en abejas
3. EFECTOS DE LA LUZ SOBRE LA PIGMENTACIÓN.
El pigmento que existe para absorber la luz o para filtrarla y tiene una función protectora (críptica) o demostrativa (fanérica) menos frecuente y de valor social o de reproducción.

Figura 12, lenguado, Función protectora de la pigmentación

Figura 13, Pavo real, función fanerica de la pigmentación
Requiere la existencia de células o grupos celulares organizados: los cromatóforos. El pigmento puede ser melanina (melanóforos), estará compuesto de carotinoides liposolubles (lipóforos) o guanina (guanóforos), siendo los dos últimos de una gran variedad cromática e iridiscentes.

Figura 14, sepia (melanóforos)

Figura 15, Camaleón (guanóforos)
La acción de la luz sobre el pigmento puede ser de tres tipos:
Directa sobre los melanóforos de la piel o incluso sobre ectodermo retiniano o iridiano: contracción pupilar directa en cefalópodos peces y anfibios.
Indirecta a través del ojo.
Indirecta a través del mesencéfalo (algunos peces) o la glándula pineal (lamprea, peces teleósteos).
4. VISIÓN PROPIAMENTE DICHA.
La función esencial y primitiva de la visión es el control de los movimientos para alcanzar tan fácilmente como sea posible un entorno óptimo. Esto se emplea para evitar obstáculos, para perseguir la presa o huir de los depredadores y en el hombre requiere una estrecha relación entre los ojos y el aparato de vestibular.

Figura 16, coordinación mano-ojo en orangután
Los niveles más altos de función visual son, por su propia naturaleza, conscientes aunque en animales inferiores (por ejemplo: la marcha de la hormiga) no lo son y tampoco, naturalmente en máquinas reguladas por mecanismos cibernéticos de retroalimentación. El proceso de “concienciación” de la visión ha coincidido, o se ha debido, a la emigración de la estación terminal desde los núcleos basales a la corteza (lo que es un hecho reciente en la historia de la evolución).
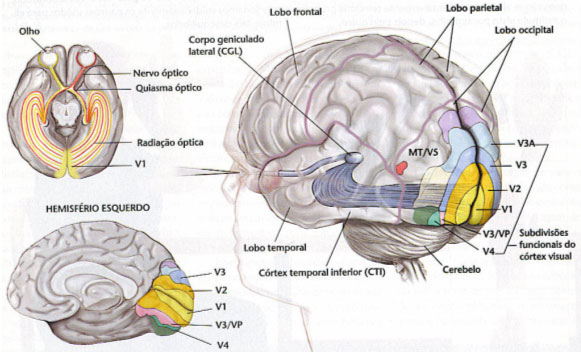
Figura 17, vías ópticas en el hombre: corticalización de la visión
LA EVOLUCIÓN DEL APARATO VISUAL
Aunque en animales inferiores hay una sensibilidad luminosa difusa en todo el ectodermo, pronto parte de este se especializa en la recepción luminosa que queda limitada a los fotoreceptores. Estas se caracterizan por la presencia de pigmentos: la melanina que es químicamente inerte y actuar sólo como absorbente y los pigmentos visuales propiamente dichos. Todos estos se basan en los carotinoides asociados o no a proteínas, que se encuentran también, además de la clorofila, en el reino vegetal (xantofila). Cuando se alcanza el nivel del ojo con capacidad óptica para formar imágenes, todos los pigmentos están relacionados concretamente con la vitamina A (A1 ó A2).
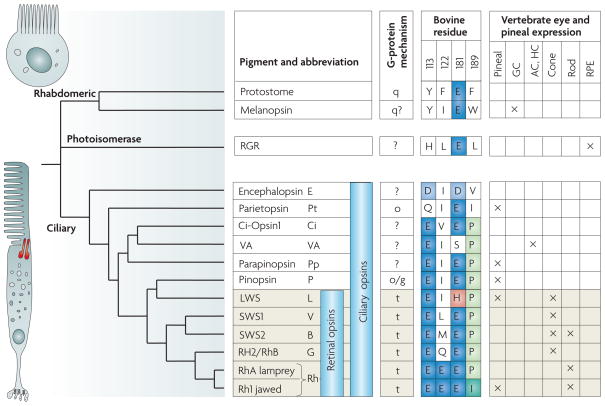
Figura 18, evolución de los fotorreceptores y los pigmentos
En los organismos unicelulares la sensibilidad luminosa difusa puede concentrarse en un punto que está asociado a los cilios o flagelos (función motora). En los flagelados este “punto ojo” o “estigma”, cuenta con pigmento y ocasionalmente con una estructura retráctil que concentra la luz y actúa, como una lente (cristalino).
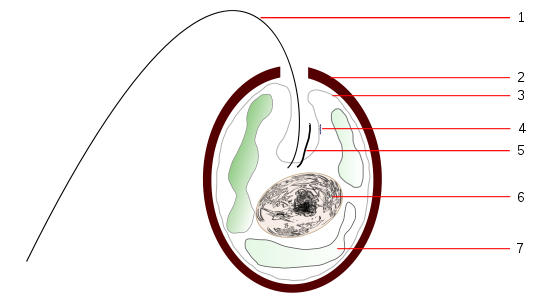
Figura 19, esquema de trachelomona
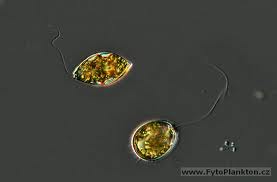
Figura 20, trachelomonas
En los seres pluricelulares pronto algunas células ectodérmicas se especializan en la recepción luminosa como células bipolares (un extremo receptor distal y otro transmisor proximal) o apolares de forma elipsoidal. Desde aquí se inicia la evolución del ojo en los invertebrados.
EL OJO DE LOS INVERTEBRADOS
Deriva del ectodermo de superficie y secundariamente conecta con el sistema nervioso.
Puede ser simple, ya sea unicelular o multicelular, o compuesto.
- Ojo simple un ocelo.
Consiste en una o varias células sensibles a la luz que actúan sin una asociación funcional.
Cuando es multicelular puede ser todavía subepitelial (sólo una acumulación de células fotosensibles) o en una fase más avanzada epitelial invaginado. Este último tipo pasa por las fases plana, cupular (conformación de una cámara oscura) y finalmente vesicular, que aún se perfecciona en el ojo de la cefalópodos en el que un engrosamiento el epitelio forma ya un “remedo” de cristalino, y hay también una pupila contráctil.

Figura 21, stilaria lacustris
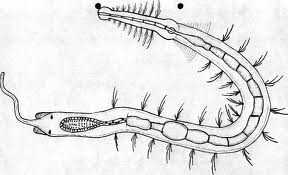
Figura 22, Anatomia Stilaria L
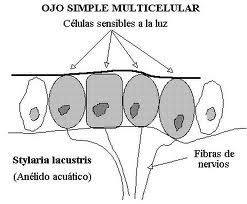
Figura 23, ojo plano, stilaria L.

Figura 24, patella sp.
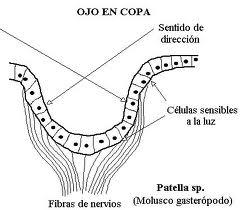
Figura 25, ojo en copa en patella SP

Figura 26, helix pomatia
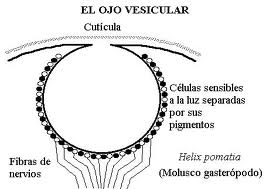
Figura 27, ojo vesicular en helix P.
Aunque la existencia de una retina invertida (en la que los elementos fotoreceptores están en la capa más profunda y los transmisores en la superficie, lo que permite una mejor nutrición de los primeros) es característica de los vertebrados, algunos moluscos y arácnidos posen ya esta nueva organización retiniana, en vez de la retina “vertida” o directa. Este cambio en la orientación de la retina es uno de los procesos de más difícil explicación a la luz de la teoría de evolución continúa. Además en animales que han de vivir en pobres condiciones de iluminación, aparece detrás de la retina una capa cristalina reflectante o tapetum.
- Ojo compuesto.
En el ojo compuesto los elementos sensoriales están agrupados estructural y sensorialmente. Cada uno de ellos, llamado omatidio, consta de una faceta corneal, un cono cristaliniano, una célula pigmentada iridiana y un retínulo compuesto por los fotoreceptores que acaban en una prolongación común a varios de ellos, llamada rabdoma, en la que se inicia la transmisión de los impulsos hacia los centros nerviosos.
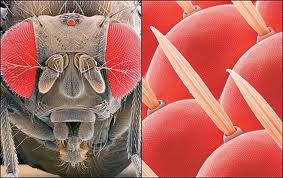
Figura 28, ojo compuesto
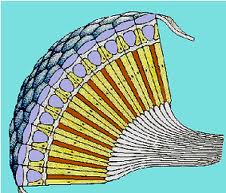
Figura 29, esquema ojo compuesto
El ojo compuesto, peculiar de los artrópodos, adopta diferentes variedades en arácnidos, crustáceos e insectos. Aunque sólo permite alcanzar una agudeza visual reducida, variable según haya una mayor o menor concentración del retínulo sobre los rabdomas, el ojo compuesto tiene la capacidad de analizar el plano de polarización de la luz, función esencial para la orientación en el espacio.
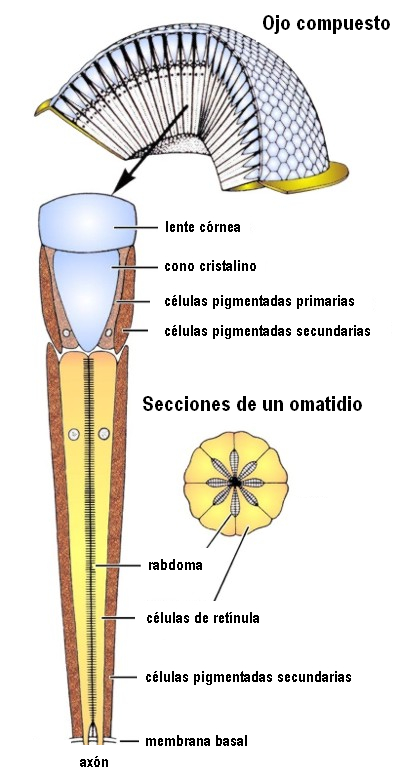
Figura 30, estructura de un omatidio
EL OJO DE LOS VERTEBRADOS
Supone una revolución evolutiva ya que, con las diferencias que veremos, tiene un patrón común en todas las especies y no se asiste aquí el proceso continuo de perfeccionamiento, a lo largo de distintas vías, que se aprecia en los invertebrados. A diferencia de los invertebrados deriva del ectodermo neural.
El ojo de los vertebrados no muestra la diferenciación creciente que se observa en el cerebro, el oído o el corazón, y el ojo de un pez es, esencialmente tan complejo y completamente desarrollado como el de un ave o mamífero. Este es sólo uno, aunque para nosotros el más interesante, de los misterios que sigue arrojando la comprensión de los mecanismos evolutivos desde los animales sin un esqueleto óseo a otros que lo poseen tan completo.
Esquemáticamente el ojo de los vertebrados se diferencia en tres tipos según el hábitat:
- El de los ictiopsidos, peces y anfibios, adaptados a un medio acuático, en los segundos sólo en la fase larvaria.
- El de los Sauropsidos, reptiles y aves, adaptados a un medio aéreo.
- El de los mamíferos, ambivalente.
El elemento fundamental, la retina, tiene la misma estructura en todos los vertebrados, aunque varíe según la iluminación la relación entre conos y bastones. Hay quien piensa que los conos son la célula visual primitiva y que los bastones son un elemento posterior debido a una transmutación relacionada con el desarrollo de la rodopsina, lo que ocurrió en primer lugar en los peces de aguas profundas. En el resto de las estructuras oculares existen en los vertebrados diferencias considerables que son la consecuencia de la adaptación a ambientes muy diversos: inicialmente el ojo se adaptó a aguas poco profundas pero después ha evolucionado, según los casos, a la existencia en aguas abismales, en los ríos, en el lodo de los pantanos, en la tierra y en el aire así como durante la noche, el crepúsculo o la semioscuridad de las cavernas o la más intensa luz diurna. Además ha alcanzado una visión panorámica cuando es esencial la percepción del movimiento o una visión estereoscópica cuando es un extracto cálculo de las distancias, oscilando entre una simple vaga apercepción y la más exacta capacidad de resolución o la más compleja visión cromática.
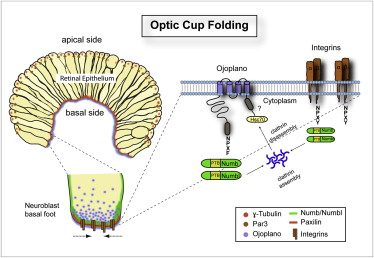
Figura 31, control del plegamiento de la cúpula óptica
Prescindiendo de los poco numerosos ciclóstomos, podemos resumir así las principales características del ojo de los peces, anfibios, reptiles, aves y mamíferos.
a. Peces.
La córnea carece prácticamente de valor dióptrico, de modo que el globo ocular tiende a ser muy grande y cristalino refringente. La córnea es aplanada para resistir mejor la presión del agua. El tamaño del globo ocular es mayor cuanto mayor lo es la profundidad de las aguas, de tal forma que en los espacios abisales, desprovistos de luz, el ojo degenera y casi desaparece. Incluso en especies como la anguila el ojo crece al desplazarse el animal desde los días de Europa o América hasta el mar de los sargazos en el atlántico donde procrean y mueren. La retina muy rica en bastones suele ser avascular. Los movimientos oculares son muy restringidos y la fijación se consigue preferentemente con movimientos de todo cuerpo y la cabeza. Ya conocemos la curiosa diferencia en los pigmentos visuales, que los peces de agua dulce están constituidos a base de vitamina A2 y en los que, como los salmones, emigran de los mares a los ríos y cambia consiguientemente el pigmento visual.
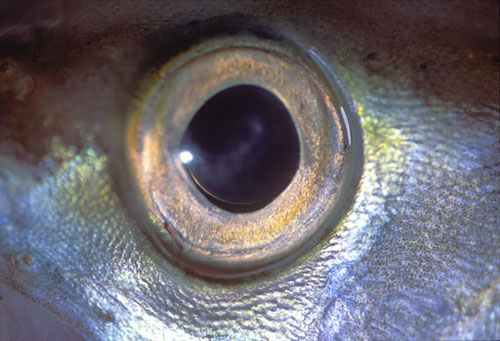
Figura 32, ojo de pez
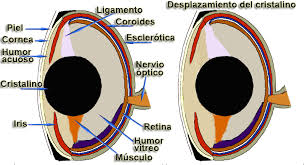
Figura 33, esquema del ojo de los peces
b. Anfibios
El paso del medio acuático al aéreo permite que la córnea adopte funciones dióptricas, haciéndose fuertemente convexa. El cristalino menos grande que los peces, permite una acomodación mediante desplazamientos antero-posteriores. Aparecen estructuras protectoras como los párpados, glándulas y vías lagrimales.

Figura 34, ojo de rana comun
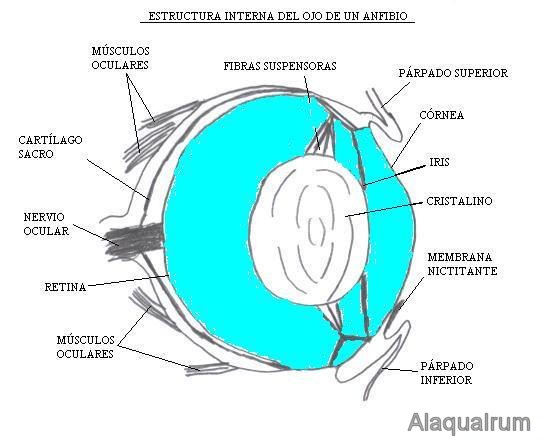
Figura 35, esquema del ojo de un anfibio
c. Reptiles.
La acomodación se lleva a cabo por medio de una deformación del cristalino, lo que requiere la existencia de un músculo ciliar que se inserta en la periferia de la córnea. La retina es avascular y se nutre a expensas de la coroides o de una membrana avascular extraretiniana. Poseen un tercer párpado.
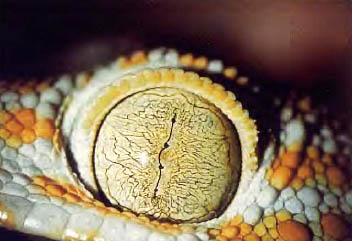
Figura 36, ojo de reptil
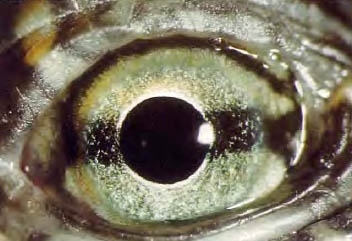
Figura 37, ojo de tortuga marina
d. Aves.
El ojo suele ser de mayor tamaño que el que aparenta con un mecanismo acomodativo por medio de deformación del cristalino. La retina es gruesa, con gran abundancia de fotoreceptores, conos y o bastones más abundantes según los hábitos de vida. La retina esta nutrida por un Pecten, membrana vascular alargada que corresponde a la papila óptica. En algunos tipos existen dos fóveas, una para visión lejana y otra para visión próxima.
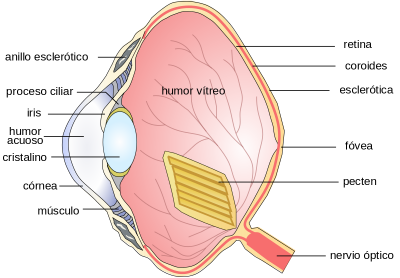
Figura 38, esquema anatómico delojo de las aves

Figura 39, ojo de buho
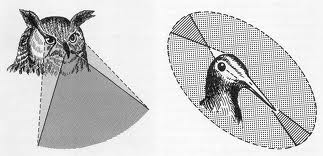
Figura 40, campo visual en aves
e. Mamíferos.
El ojo de los monotremas, ornitorrinco, es muy parecido al de los reptiles, el de los marsupiales tiene una estructura intermedia entre el de los reptiles y el de los placentados. Esta es la subclase más abundante y variada y comprenden quince órdenes: insectívoros, quirópteros, dermatópteros, primates (con varios subórdenes y familias), roedores, carnívoros, artiodáctilos, perisodáctilos, proboscídeos, sirénidos, cetáceos…
El ojo de los placentados difiere del resto de los vertebrados en tres aspectos fundamentales:
- El desarrollo y posterior involución de la arteria hialoidea, que en muchos casos da lugar a una vascularización intraretiniana.
- En la aparición de la capa mesodérmica del iris.
- En el mecanismo acomodativo debido a una deformación de la cápsula del cristalino.
Como es natural el hábitat y los hábitos de vida imponen variaciones menores en el resto de las estructuras oculares: cristalino casi esférico en los que viven en un medio acuático, presencia o ausencia de un tapetum situado entre la coroides y la retina, las variaciones en el grosor de la esclera (que en la ballena es de3/4 del diámetro ocular), en la forma de la pupila y del esfínter, en la vascularización de la retina (total, parcial o ausente: retinas holoangióticas, mesoangióticas y anangióticas) y en la proporción de conos y bastones retinianos.
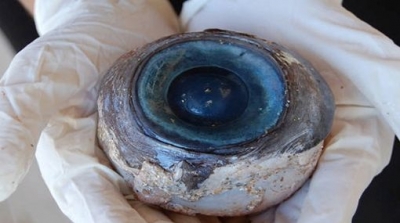
Figura 41, ojo de ballena
Uno de los aspectos más interesantes (ya que la descripción detallada de cada estructura ocular en cada orden, suborden y familia nos llevaría demasiado lejos) es el de la decusación de las fibras ópticas.
En los demás vertebrados la decusación es prácticamente total, lo que permite un campo visual binocular que puede abarcar los 360° pero limite a la visión estereoscópica a mecanismos menos exactos que los mamíferos. En estos, sobre todo en los placentados, empieza a ver fibras directas en relación a la frontalización de los ojos. La proporción es de 1/6 del total en el caballo, 1/4 a 1/3 en el perro y al gato, 1/3 en los primates superiores y casi 1/2 en el hombre. Este proceso supone una pérdida de la extensión del campo visual binocular pero la adquisición de una exacta visión estereoscópica. Se ha especulado en que ésta empezó a ser necesaria en animales que vivían en un medio arbóreo, en el cual los desplazamientos derramar a rama exigen un adecuado cálculo de las distancias y se asoció al desarrollo de la mano como órgano prensil.
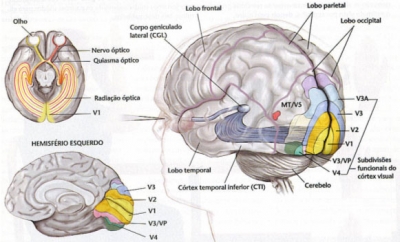
Figura 42, decusación de las vías visuales en el hombre
Ya hemos señalado antes que sólo en los mamíferos aparece una zona del córtex cerebral a la que llegan y en la que son analizados los impulsos visuales. Aun así, excepto en el hombre, está localización es aún imprecisa y en la mayor parte de los mamíferos la ablación del córtex visual no ocasionar una ceguera completa. En el hombre en cambio la única actividad visual que sigue siendo subcortical es la pupilar.
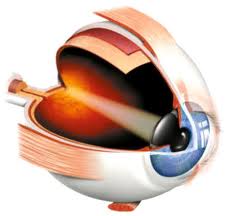
Figura 43, ojo humano, esquema
FUNCIÓN VISUAL
El medio ambiente y los hábitos de vida, preferentemente la actividad diurna o nocturna, imponen una amplia gama de variaciones en todos los aspectos de la función visual: percepción luminosa, sentido cromático, sentido de la forman, visión estereoscópica…
Sirva de ejemplo un resumen de los distintos mecanismos mediante los cuales se consigue una acomodación en los vertebrados:
Mecanismos estáticos:
Una pupila estenopéica.
Un sistema óptico duplicado.
La interposición de una membrana nictitante.
Una retina duplicada.
Una retina en rampa.
Una retina rugosa.
Una gran longitud de los elementos receptores.
Mecanismos dinámicos:
Desplazamiento de cristalino hacia adelante o hacia atrás, inducidos por presión corneal o por una musculatura propia.
Deformación de cristalino, que los mamíferos es debido en último extremo a la elasticidad de la cápsula y a su distinto grosor.
La agudeza visual llega en las aves alcanzar una resolución de 10” de arco, tres veces superior a la del hombre.
El campo visual monocular oscila entre los 30° del camaleón y los 215 grados del caballo. En los peces sumergidos se conjuga la visión acuática y la aérea, pero sometida está a la restricción que impone la refracción total de la superficie del agua. El binocular como señalábamos antes, abarca los 360° en los animales que pueden ser presas y se reduce en los depredadores. En los peces con ojos de posición lateral el área ciega, considerando una superficie esférica puede llegar a ser muy reducida.
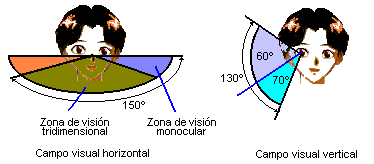
Figura 44, campo visual en el hombre
Al mismo tiempo hay una gran variedad con el ángulo que forman los ejes ópticos y la línea media, al igual que en el límite de la motilidad ocular sólo conjugada en los mamíferos.
5. ÍNDICE DE FIGURAS.
- Figura 1, luz y metabolismo de las plantas.
- Figura 2, Ciclo sexual femenino.
- Figura 3, fototropismo vegetal en girasoles.
- Figura 4, mecanismo fototropismo (auxina).
- Figura 5, clinoquinesis en la ameba.
- Figura 6, fototaxis en polillas.
- Figura 7, clinotaxis en euglena.
- Figura 8, tropotaxis en gusano.
- Figura 9, ojo compuesto.
- Figura 10, menotaxis en aves.
- Figura 11, mnemotaxis en abejas.
- Figura 12, lenguado, Función protectora de la pigmentación.
- Figura 13, Pavo real, función fanerica de la pigmentación.
- Figura 14, sepia (melanóforos).
- Figura 15, Camaleón (guanóforos).
- Figura 16, coordinación mano-ojo en orangután.
- Figura 17, vías ópticas en el hombre: corticalización de la visión.
- Figura 18, evolución de los fotorreceptores y los pigmentos.
- Figura 19, esquema de trachelomona.
- Figura 20, trachelomonas.
- Figura 21, stilaria lacustris.
- Figura 22, anatomia Stilaria L.
- Figura 23, ojo plano, stilaria L.
- Figura 24, patella sp.
- Figura 25, ojo en copa en patella SP.
- Figura 26, helix pomatia.
- Figura 27, ojo vesicular en helix P.
- Figura 28, ojo compuesto.
- Figura 29, esquema ojo compuesto.
- Figura 30, estructura de un omatidio.
- Figura 31, control delplegamiento de la cúpula óptica.
- Figura 32, ojo de pez.
- Figura 33, esque del ojo de los peces.
- Figura 34, ojo de rana comun.
- Figura 35, esquema del ojo de un anfibio.
- Figura 36, ojo de reptil.
- Figura 37, ojo de tortuga marina.
- Figura 38, esquema anatómico delojo de las aves.
- Figura 39, ojo de buho.
- Figura 40, campo visual en aves.
- Figura 41, ojo de ballena.
- Figura 42, decusación de las vías visuales en el hombre.
- Figura 43, ojo humano, esquema.
- Figura 44, campo visual en el hombre.
6. BIBLIOGRAFIA.
- The eye in evolution, System of Ophthalmology Volumen I, Sir Stewart Duke-Elder, The C.V. Mosby Company, 1958.
- Animal Eyes, Michael F. Land, Dan-Eric Nilsson, 2ª Edition, 2012, Oxford Animal Biology Series.
- Webvision, The Organization of the Retina and Visual System, http://webvision.med.utah.edu/
- The origen of colour visión in vertebrates, Shaun P Collin and Ann EO Trezise, Clinical and experimental optometry, 2004.
- The 'division of labour' model of eye evolution, Detlev Arendt, Harald Hausen and Günter Purschke, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0104, published 31 August 2009.
- The evolution of early vertebrate photoreceptors, Shaun P. Collin, Wayne L. Davies, Nathan S. Hart and David M. Hunt, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0099, published 31 August 2009.
- Evolution and spectral tuning of visual pigments in birds and mammals, David M. Hunt, Livia S. Carvalho, Jill A. Cowing and Wayne L. Davies, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0044, published 31 August 2009.
- Evolution of colour vision in mammals, Gerald H. Jacobs, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0039, published 31 August 2009.
- Evolution of phototaxis, Gáspár Jékely, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0072, published 31 August 2009.
- Evolution and the origin of the visual retinoid cycle in vertebrates, Takehiro G. Kusakabe, Noriko Takimoto, Minghao Jin and Motoyuki Tsuda, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0043, published 31 August 2009.
- The evolution of phototransduction and eyes, Trevor D. Lamb, Detlev Arendt and Shaun P. Collin, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0106, published 31 August 2009.
- Evolution of vertebrate retinal photoreception, Trevor D. Lamb, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0102, published 31 August 2009.
- The evolution of eyes and visually guided behaviour, Dan-Eric Nilsson, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0083, published 31 August 2009.
- The evolution of irradiance detection: melanopsin and the non-visual opsins, Stuart N. Peirson, Stephanie Halford and Russell G. Foster, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0050, published 31 August 2009.
- Evolution of opsins and phototransduction, Yoshinori Shichida and Take Matsuyama, Phil. Trans. R. Soc. B 2009 364, doi: 10.1098/rstb.2009.0051, published 31 August 2009.